Norovirus Structure, Transmission, and Latest Research Advances: From Molecular Mechanisms to Vaccine Development
Release date:
2026-05-14 View count: 1
On April 28, 2026, the American Princess Cruises ship "Caribbean Princess" experienced a norovirus outbreak. The ship departed from Fort Lauderdale, Florida, and gastrointestinal illness began spreading during the voyage. According to data from the U.S. Centers for Disease Control and Prevention (CDC), a total of 102 passengers and 13 crew members had reported symptoms by the time of the report, primarily diarrhea and vomiting. The infection rate was approximately 3.3% among 3,116 passengers and 1.2% among 1,131 crew members. The CDC confirmed norovirus as the causative pathogen and recommended enhanced disinfection, case isolation, and sample testing. This incident once again highlights the ease with which norovirus can cause clustered outbreaks in high-density, enclosed environments. Shared facilities, food, and frequent surface contact among passengers provide ideal conditions for viral transmission.
Figure 1. American Princess Cruise Ship (Image from the internet)
Norovirus (NoV) is a member of the Caliciviridae family, genus Norovirus, and is one of the leading causes of acute gastroenteritis worldwide. It is estimated to cause hundreds of millions of cases of acute gastroenteritis each year. The virus can lead to more severe dehydration and complications in children, the elderly, and immunocompromised individuals. Based on genetic characteristics, noroviruses are divided into multiple genogroups and genotypes, with GI, GII, GIV, GVIII, and GIX being the main ones associated with human infections. GII.4 has been the dominant genotype globally for the past two decades, and its antigenic diversity is closely linked to its epidemic characteristics.
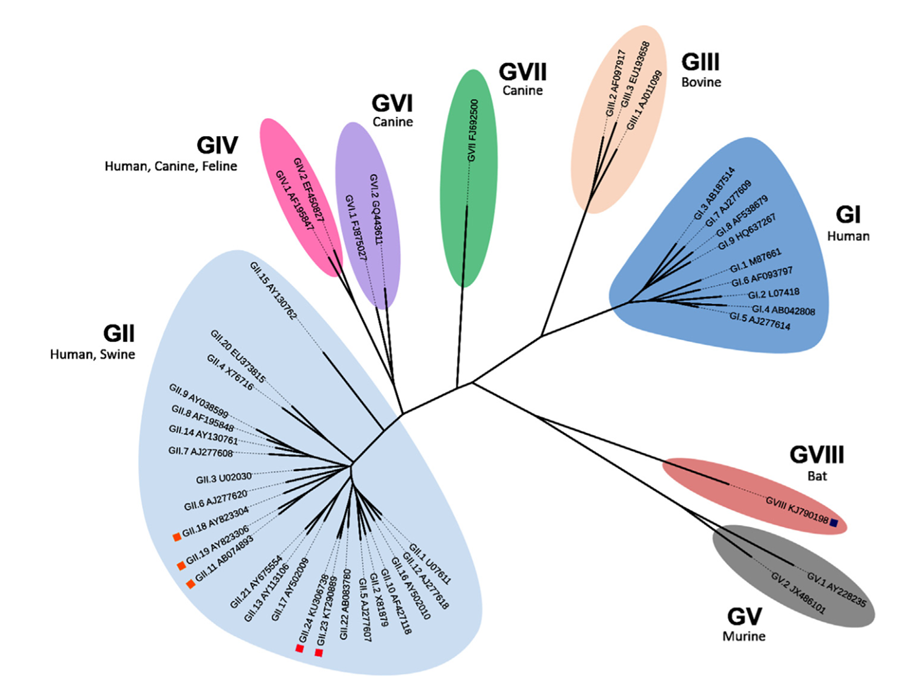
Figure 2. Phylogenetic analysis of complete VP1 amino acid sequences divides noroviruses into eight genogroups (GGI-GVII) and 44 genotypes (doi: 10.1016/B978-0-12-804177-2.00004-X)
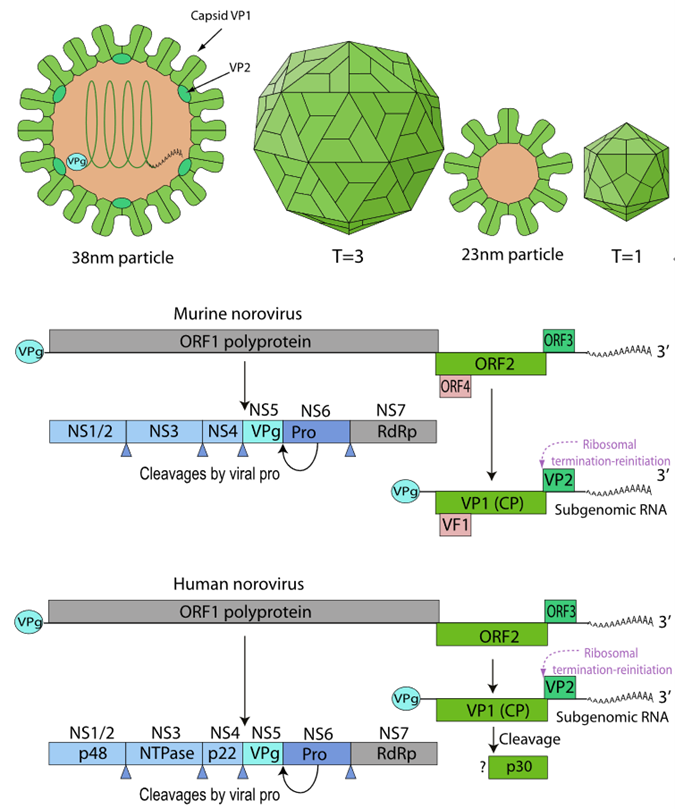
The viral particles are non-enveloped and icosahedral, with a diameter of approximately 27–40 nm. The genome is a single-stranded positive-sense RNA of about 7.5–7.7 kb in length, with a VPg (viral genome-linked protein) covalently attached to the 5' end and a polyadenylated 3' end. The genome contains three major open reading frames (ORFs): ORF1 encodes a non-structural polyprotein, ORF2 encodes the major capsid protein VP1, and ORF3 encodes the minor capsid protein VP2.
Figure 3. Virus structure and genome organization
Molecular Structure and Protein Functions
Non-structural Proteins
The polyprotein encoded by ORF1 is processed by the viral 3C-like protease (NS6) into six non-structural proteins (NS1/2 to NS7), which are involved in RNA replication, membrane rearrangement, and host regulation.
Protein Name
Common Name
Core Function and Molecular Mechanism
NS1/2
p48
Immune Regulation and Secretion Inhibition: Regulates RNA-dependent RNA polymerase (RdRp) activity and influences host immune and secretory pathways.
NS3
NTPase (p41)
RNA Translocation and Membrane Rearrangement: Possesses nucleoside triphosphatase activity, belongs to the AAA+ ATPase family, often exists as a hexamer; involved in RNA unwinding and membrane reorganization around the viral replication complex (RC).
NS4
p22
Organelle Function Disruption: Induces host cell membrane rearrangement by mimicking COPII vesicle export signals; affects Golgi apparatus function.
NS5
VPg
Translation Initiation and Genome Protection: Covalently binds to the 5' end of viral RNA; interacts with host translation initiation factors (eIFs) to direct viral protein synthesis.
NS6
Protease (Pro)
Polyprotein Processing: A cysteine protease responsible for precisely cleaving the long polyprotein encoded by ORF1 into individual functional non-structural protein units.
NS7
RdRp
Genome Replication Core: RNA-dependent RNA polymerase, the core component of the viral replication machinery, directly responsible for synthesizing both positive- and negative-strand viral RNA.
Structural Proteins
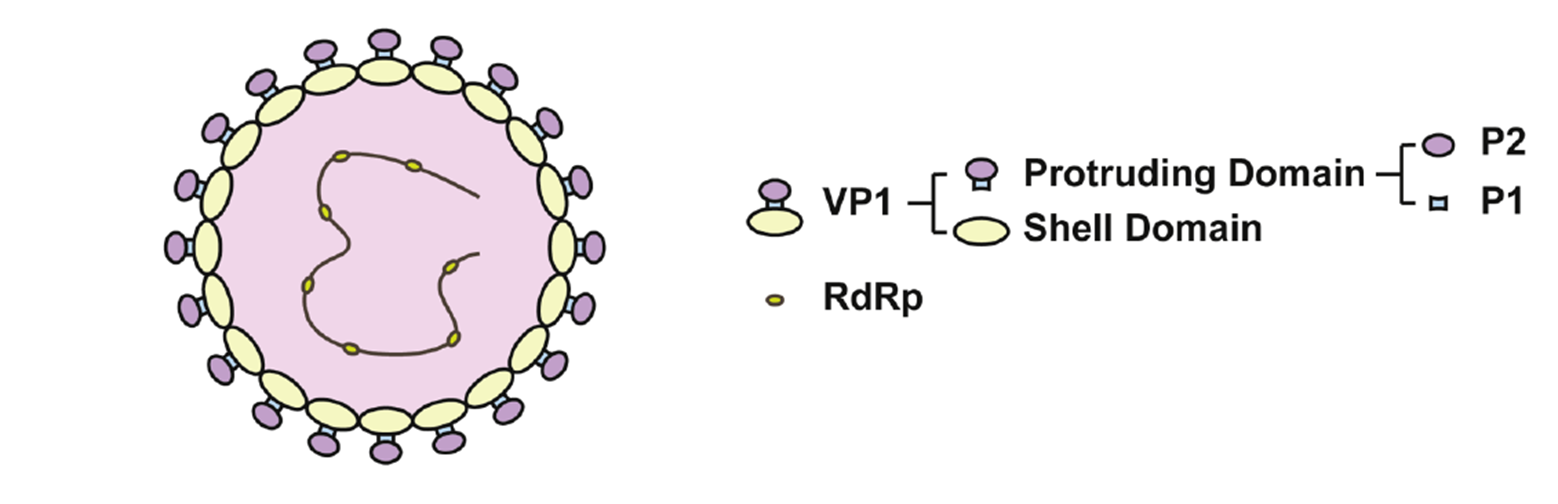
VP1 (Major capsid protein, encoded by ORF2): Forms 90 dimers that make up the viral capsid and is critical for viral attachment, immunogenicity, and structural stability. VP1 can self-assemble into virus-like particles (VLPs). Its P domain contains receptor-binding sites (such as binding to histo-blood group antigens, HBGAs). VP1 is the primary site of antigenic variation and a key target for vaccine and diagnostic development.
Figure 4. VP1 structural organization: S domain (shell) and P domain (protrusion); the P2 subdomain mediates interaction with host HBGAs
VP2 (Minor capsid protein, encoded by ORF3): Located inside the capsid and involved in RNA packaging and capsid stability. The virus is non-enveloped and highly environmentally stable, surviving on surfaces for days and showing some resistance to certain disinfectants — characteristics closely related to its capsid structure.
Latest Research Advances
Norovirus is the primary non-bacterial pathogen responsible for global epidemics of acute gastroenteritis (AGE). Due to its high genetic heterogeneity and the lack of robust in vitro culture systems, specific prevention methods have long been unavailable. Currently, there are no approved norovirus vaccines or specific antiviral drugs. Research efforts mainly focus on VP1-based strategies.
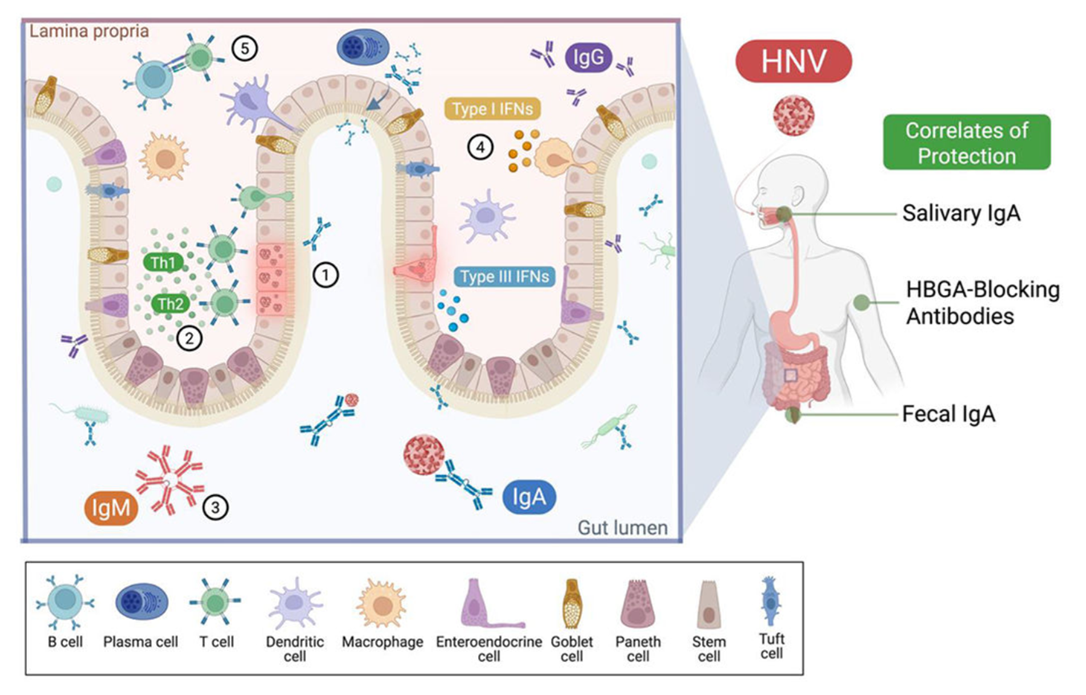
Figure 5. Working model of HNV immune interactions and protection
1. Vaccine Development: Transition from VLPs to Multivalent mRNA Platforms
Although virus-like particle (VLP) vaccines based on VP1 protein remain the mainstream approach, their lag in responding to antigenic drift has prompted researchers to shift toward more flexible mRNA platforms. Latest data from 2025–2026 experiments show that lipid nanoparticle (LNP) vaccines encapsulating VP1 sequences from multiple genotypes (GI.1, GII.4, GII.2, GII.17, GIX.1) induced broader-spectrum neutralizing antibody titers in animal models. Compared with traditional VLPs, mRNA vaccines significantly activate Th1-type T cell responses, which are crucial for clearing intracellular infections and establishing long-term immune memory.
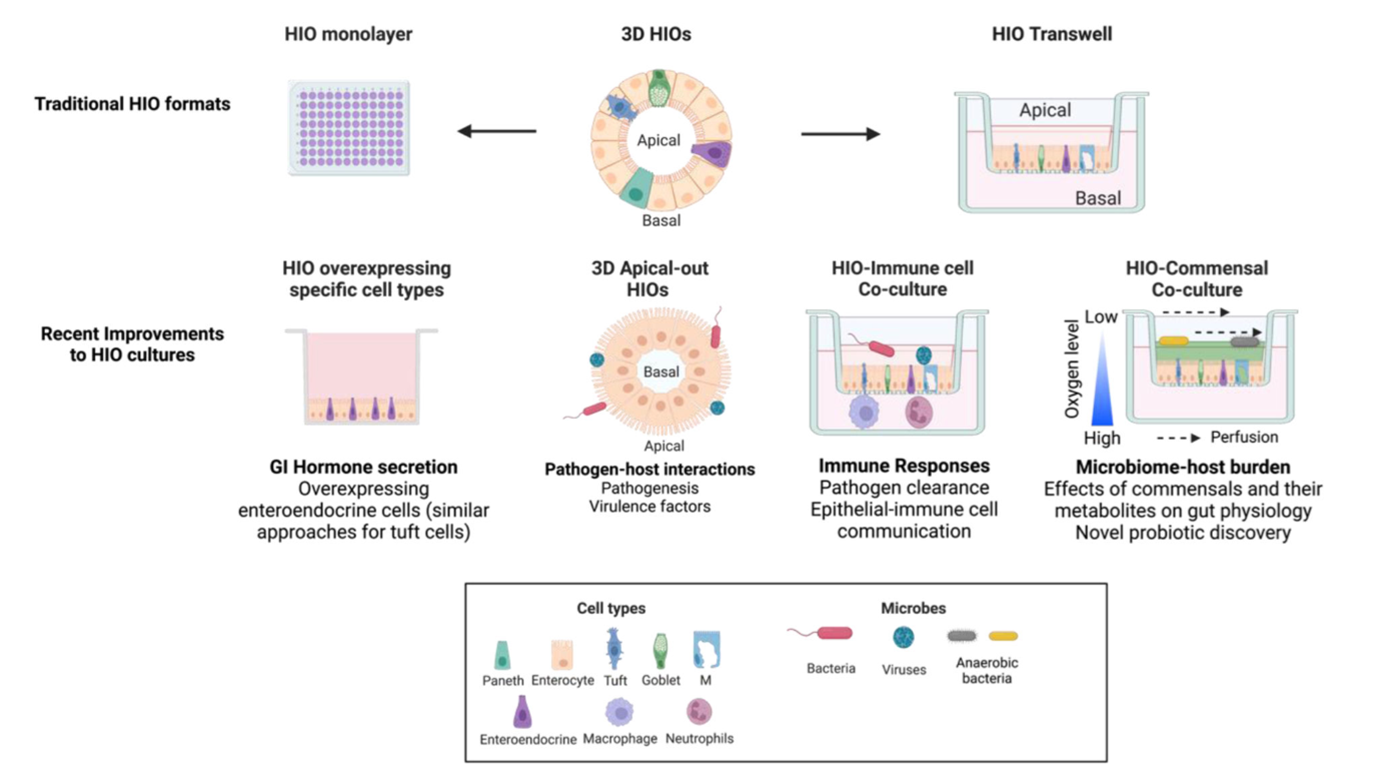
2. In Vitro Models: Human Intestinal Organoids (HIOs) Reveal Infection Mechanisms
For a long time, norovirus research has been limited by the failure of traditional cell line cultures. The establishment of human intestinal organoids (HIOs) has provided a highly faithful in vitro platform for studying viral replication mechanisms and drug screening. Recent studies have confirmed that invasion by certain GII strains depends not only on host cell surface histo-blood group antigens (HBGAs) but also requires bile salts as cofactors to induce conformational changes in the viral capsid, thereby promoting endocytosis. Using CRISPR-Cas9 to edit the FUT2 gene in organoids, researchers have further clarified the differential susceptibility of secretor and non-secretor populations to different norovirus genotypes.
Figure 6. Tissue-derived organoid cultures used to study intestinal bacteria and viruses
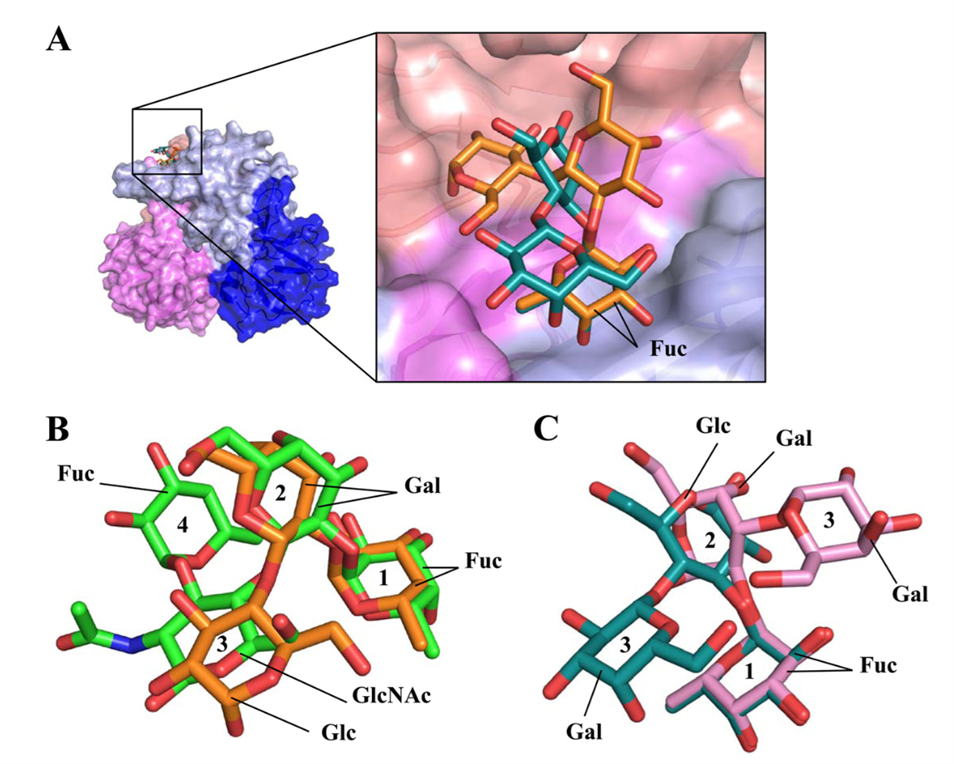
3. Antiviral Strategies: Competitive Inhibition by Human Milk Oligosaccharides (HMOs)
In the field of non-vaccine prevention, strategies targeting viral attachment receptors have made significant progress. Studies have shown that human milk oligosaccharides (HMOs), particularly fucosylated components, have spatial configurations highly similar to HBGAs on intestinal epithelial cells. Structural biology research indicates that HMOs can precisely bind to the P domain of VP1, competitively occupying binding sites and blocking viral attachment to host cells.
Figure 7. Binding of HMOs and HBGAs to GII.10 norovirus
4. Key Antigenic Target: Advanced Structural and Functional Studies of VP1 Protein
VP1, the major structural protein of norovirus, consists of an S (shell) domain and a P (protruding) domain, with the P domain determining viral antigenicity and receptor-binding specificity. Recent studies using the insect cell-baculovirus expression vector system (IC-BEVS) have achieved high-purity self-assembly of P particles, laying the foundation for high-throughput epitope mapping and diagnostic kit development. Other research is using single-cell B cell sequencing to screen high-potency neutralizing antibodies targeting conserved regions of the P domain from recovered patients, providing candidate drugs for emergency immunotherapy.
Prevention and Control Significance
Norovirus’s characteristics of low infectious dose, high environmental stability, and efficient person-to-person transmission make it prone to causing clustered outbreaks in enclosed or semi-enclosed settings. Prevention and control rely on strict hygiene measures and food safety management. Advances in vaccine development are expected to provide additional protection tools for high-risk environments. With further progress in culture systems, structural biology, and immunology research, intervention measures against norovirus will gradually improve.
Campillax-Veliz CP, Carvajal J, Avellaneda AM, Escobar D. Covian C. Kalergis AM.Lay MK. Human Norovirus Proteins: Implications in the Replicative Cycle,Pathogenesis, and the Host Immune Response. Front Immunol. 2020 Jun 16:11:961.doi: 10.3389/fimmu.2020.00961. PMID: 32612600; PMCID: PMC7308418.
Flitter BA, Gillard J, Greco SN, Apkarian MD, D'Amato NP, Nguyen LQ. Neuhaus ED,Hailey DCM. Pasetti MF, Shriver M. Quigley C. Erenck RW Jr. Lindesmith LC, Baric RS,Wei LJ, Tucker SN, Cummings JF. An oral norovirus vaccine generates mucosalimmunity and reduces viral shedding in a phase 2 placebo-controlled challengestudy. Sci Transl Med. 2025 May 14;17(798):eadh9906. doi:10.1126/scitranslmed.adh9906. Epub 2025 May 14. PMID: 40367195.
Hou W. Ly L, Wang Y, Xing M, Guo Y, Xie D, Wei X, Zhang X, Liu H, Ren J, Zhou D36-Valent Virus-Like Particle-Based Vaccine Induced Potent and Sustained ImmunityAgainst Noroviruses in Mice. Front Immunol. 2022 May 23:13:906275. doj10.3389/fimmu.2022.906275. PMID: 35711416; PMCID: PMC9197435.
Morrin ST, Buck RH, Hill DR. A unique blend of five human milk oligosaccharidessupports recovery of infant microbiome composition and function after exwivo antibiotic use. Front Pediat 2026 Mar 5:14:1765159. doj10.3389/fped.2026.1765159. PMID: 41867919; PMCID: PMC12999568.
Wei J, Bystol K, Alabanza L, Lim S, Greig J, Gopal P, Reyes Ballista JM, Galli JD.Warren C, Swartz A, David C, Jeong U, Gonzalez-Fernandez E, DeWitt LA, Go G,Lago A, Radcliffe S, May Fulton CD, Shen P. Fridman A, Gaspar J, Pirrone A, Wen Z.O'Donnell G, Austin L, McKenney D. Krosky P, Liu Y, Bett AJ, Zhang L.Characterization of humoral and cellular immunity induced by mRNA vaccinesexpressing norovirus VP1 proteins in mice and NHPs. Mol Ther, 2026 Jan7;34(1):300-314. doi: 10.1016/j.ymthe.2025.09.023. Epub 2025 Sep 12. PMID:40946165; PMCID: PMC12925784.
Adeniyi-lpadeola G. Nwanosike H, Ramani S. Human intestinal organoids as modelsto study enteric bacteria and viruses. Curr Opin Microbiol 2023 Oct:75:102362. doi:10.1016/j.mib.2023.102362. Epub 2023 Aug 1. PMID: 37536261; PMCID:PMC10529792.e
Okten AB, Craft JE, Wilen CB. Mechanisms of Norovirus Immunity: Implications forVaccine Design. Annu Rev Pathol 2026 Jan;21(1):295-319. doj 10.1146/annurev-pathmechdis-042524-021922. Epub 2025 Oct 16. PMID: 41100711; PMCID:PMC13020347.
Zhang Q, Zhu S, Zhang X, Su L, Ni J, Zhang Y, Fang L. Recent insights into reversegenetics of norovirus. Virus Res. 2023 Feb:325:199046. doi10.1016/j.virusres.2023.199046. Epub 2023 Jan 16. PMID: 36657615; PMCID:PMC10194155.e
Weichert S. Koromyslova A, Singh BK, Hansman S. Jennewein S. Schroten H.Hansman GS. Structural Basis for Norovirus Inhibition by Human MilkOligosaccharides. J Virol 2016 Apr 14:90(9):4843-4848. doi: 10.1128/JVI.03223-15.PMID: 26889023; PMCID: PMC4836343.

 中文
中文 English
English 한국어
한국어 日本語
日本語 Español
Español Français
Français Русский
Русский