Analysis of China's First Imported South African Type 1 (SAT1) Foot-and-Mouth Disease Outbreak: Virus Structure, Antigen Differences, and Emergency Vaccine Control Strategies
Recently, China reported its first imported South African Type 1 (SAT1) foot-and-mouth disease outbreak. This serotype is an African-specific strain that differs fundamentally from China's long-circulating O, A, and Asia 1 strains in genome organization, capsid structure, antigen protein composition, and epitope conformation. There is no cross-protective immunity between different serotypes, and existing routine vaccines cannot provide effective protection.
FMDV Molecular Structure
Foot-and-mouth disease virus (FMDV) belongs to the genus Aphthovirus in the family Picornaviridae. It is a non-enveloped, icosahedral virus approximately 25–30 nm in diameter. Its genome is a single-stranded positive-sense RNA of about 8.5 kb encoding a single open reading frame, which is processed by proteases into structural and non-structural proteins. FMDV has 7 major serotypes (O, A, C, Asia 1, SAT1–3) and numerous subtypes. There is no cross-protective immunity between different serotypes. The viral RNA replication process is prone to mutations, leading to the continuous emergence of new variant strains—this is a major challenge for traditional vaccine-based control.
The SAT1 serotype shares the same genome organization as O, A, and Asia 1 strains, consisting of 5′UTR‑L‑P1‑P2‑P3‑3′UTR‑poly(A). The P1 region encodes the four structural proteins VP1, VP2, VP3, and VP4 that form the viral capsid. The P2 and P3 regions encode non-structural proteins such as Lpro, 2A, 2B, 2C, 3A, 3B, 3Cpro, and 3Dpol, which are involved in viral replication, protein processing, and immune evasion.
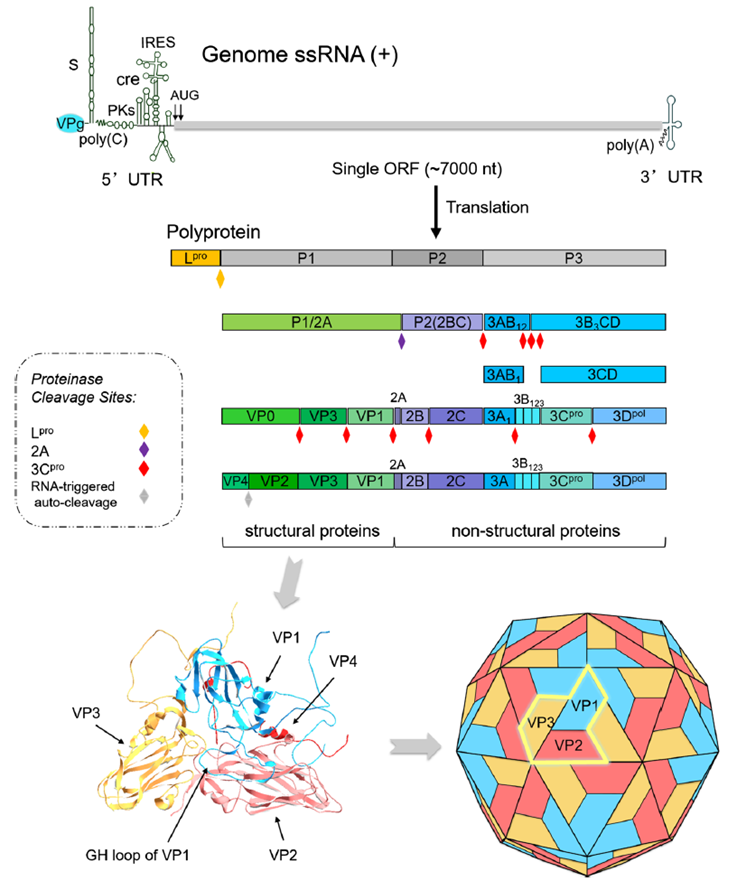
Figure 1. Foot-and-mouth disease virus (FMDV) genome, viral polyprotein processing and structural protein conformation
Among the structural proteins, VP1, VP2, and VP3 are exposed on the surface of the virion and serve as the main antigens that induce neutralizing antibodies, while VP4 is located inside the capsid and does not participate in antigen recognition. VP1 is the key protein determining serotype specificity. Its G‑H loop and C-terminus are the primary neutralizing epitope regions. The nucleotide and amino acid sequence differences in the VP1 coding region between SAT1 and common circulating strains exceed 40%, which forms the molecular basis for serotype specificity and the lack of cross-protection. The VP1 G‑H loop of common circulating strains is relatively conserved and matches well with existing vaccine antigens, inducing specific neutralizing antibodies. In contrast, the SAT1 VP1 G‑H loop is a hypervariable region with concentrated amino acid substitutions, significantly altered conformation, and unique disulfide bonds that stabilize the local structure, preventing antibodies induced by conventional vaccines from recognizing it. Although the SAT1 VP1 retains the highly conserved RGD motif, which mediates entry via host cell αvβ6 integrin, its surface loops carry additional positively charged amino acids that enable the use of auxiliary receptors such as heparan sulfate, resulting in higher cell-binding efficiency and stronger tissue tropism. VP2 and VP3 in SAT1 also contain key amino acid substitutions in regions such as the B‑C and E‑F loops, leading to remodeling of conformation-dependent antigenic epitopes and no cross-reactivity with O, A, or Asia 1 antibodies. VP4 is highly conserved across all serotypes, located inside the capsid, does not induce protective neutralizing antibodies, and shows no serotype-specific differences.
The non-structural proteins of SAT1 and common circulating strains show high overall homology, but antigenic differences exist. Lpro and 3Cpro inhibit the type I interferon pathway by cleaving host molecules such as eIF4G and RIG‑I/MDA5. 3Dpol is the RNA-dependent RNA polymerase responsible for genome replication. 3A is involved in membrane rearrangement and host range regulation. In diagnostic applications, the 3ABC antibody detection strategy can effectively differentiate naturally infected animals from those immunized with inactivated vaccines. This method is not affected by differences between SAT1 and other serotypes and is universally applicable.
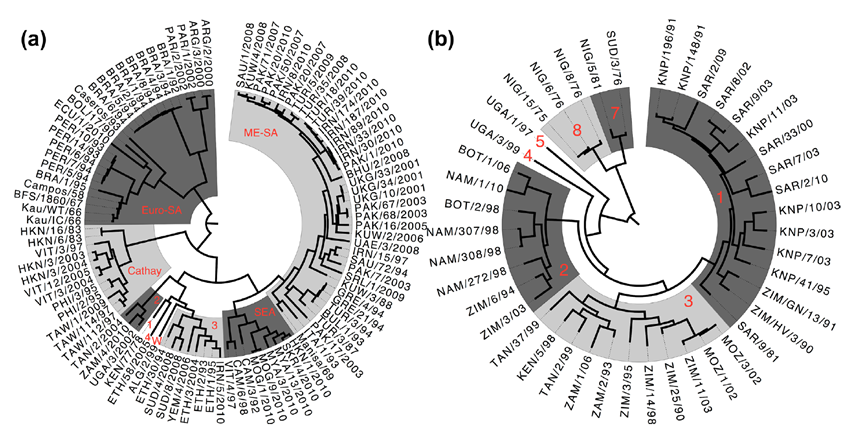
Figure 2. Phylogenetic tree of O-type and SAT1-type serogroups
From a structural and antigenic perspective, the core differences between SAT1 and common circulating strains lie in the sequence and conformation of surface antigenic epitopes on the capsid, particularly the hypervariability of the VP1 G‑H loop and the presence of unique disulfide bonds. This directly results in the inability of conventional vaccine-induced antibodies to neutralize SAT1 virus. The complete lack of cross-protective immunity between serotypes is the fundamental reason why foot-and-mouth disease control must use serotype-specific immunization and dedicated vaccines. As an exotic imported strain, SAT1 encounters a completely naïve livestock population in China. It exhibits high transmission efficiency, high mortality in young animals, and more severe clinical lesions, posing a major threat to the livestock industry.
Transmission Routes and Pathogenic Mechanisms
FMDV is one of the most contagious animal viruses known. Its basic reproduction number (R₀) in cattle herds can reach 2.52–14, and in small-scale experiments it can even show near-infinite transmission. The virus spreads primarily through direct contact, respiratory droplets, and contaminated fomites. After invading the host, it first replicates in oral mucosa and hoof epithelial cells, then disseminates systemically via the bloodstream.
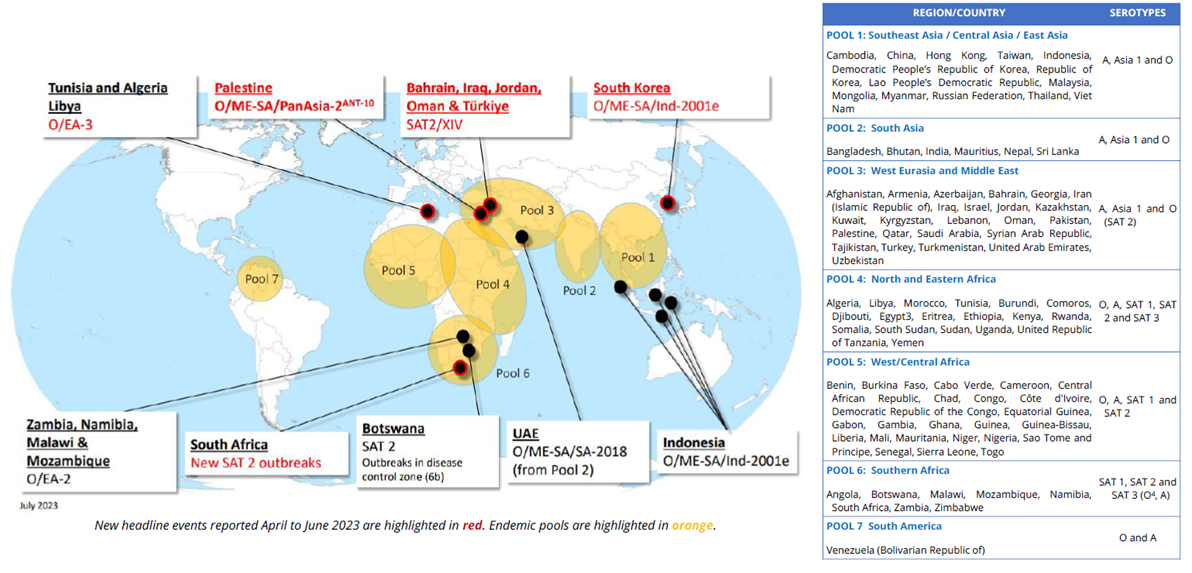
Figure 3. Geographic distribution of FMD and recent global outbreaks
During pathogenesis, the VP1 protein of FMDV binds to αvβ6 integrin on the host cell surface, mediating virus internalization. Replication leads to necrosis and shedding of epithelial cells, forming characteristic oral vesicles and hoof ulcers. Although mortality in adult animals is low, it causes significant production losses: milk yield in dairy cows drops by 20%–80%, weight gain in fattening animals decreases by 10%–25%, abortion rates in breeding animals can reach 28.8%, and loss of traction power in infected animals can persist for more than 20 days. More seriously, the virus can persist in wild animals such as African buffalo, forming natural reservoirs that pose enormous challenges to disease eradication.
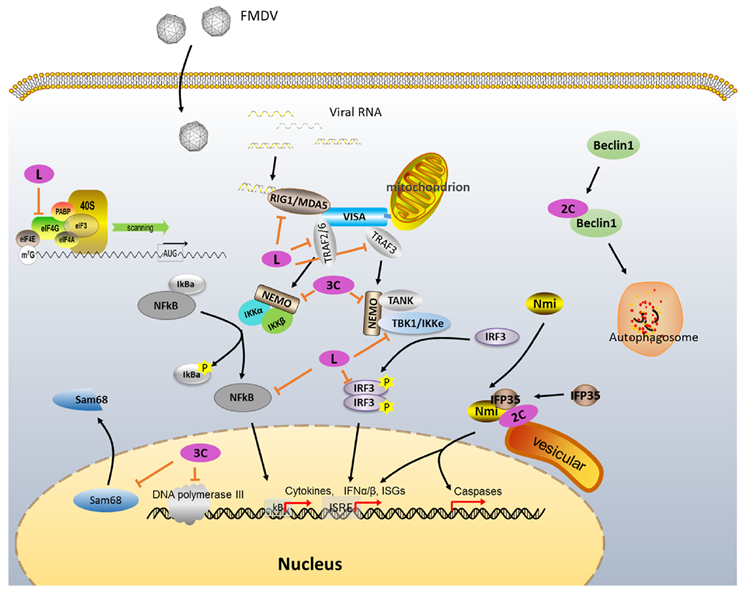
Figure 4. Known functions of FMDV non-structural proteins in cellular regulation
Current Research Progress
Structural biology and pathogen evolution studies have confirmed that the hypervariability of the SAT1-type foot-and-mouth disease virus VP1 G‑H loop, unique disulfide bond conformation, and remodeling of surface antigenic epitopes are the core molecular mechanisms underlying the lack of cross-protection with O/A/Asia1 types. Relevant findings have been published in prestigious journals such as Veterinary Research, providing critical insights for the design of precision vaccines.
Internationally, commercial vaccines against SAT1 are widely used in Africa and the Middle East, including Bioaftogen® (Biogénesis Bagó, Argentina; SAT1/SAT2 multivalent inactivated vaccine), Aftovaxpur DOE (Boehringer Ingelheim, containing SAT1/SAT2 components), and Morocco Biopharma quadrivalent vaccine (O+A+SAT1+SAT2). All have passed WOAH potency standards and can induce stable neutralizing antibodies in cattle herds, meeting regional control needs.
Following the first importation of SAT1 into China, emergency research and development was rapidly initiated. The Ministry of Agriculture and Rural Affairs approved two emergency vaccines: one inactivated vaccine (Re‑SAT1/2026 strain) produced by China Agricultural Vet. Bio. Sci. Co., Ltd. (approved 2026‑04‑01) and one subunit vaccine produced by Jinyu Baoling Bio-Pharmaceutical Co., Ltd. (a subsidiary of Jinyu Bio). Both are based on strains isolated and identified in China. Their immunogenicity and safety have been verified in clinical trials. They offer DIVA compatibility and no risk of virus shedding, and have already been deployed for emergency immunization in affected areas. In parallel, SAT1-specific fluorescent RT‑PCR and ELISA diagnostic kits have been approved, forming a precision control system combining serotype-specific detection with dedicated vaccines.
abinScience Related Products
Below is the latest catalog of abinScience recombinant proteins and antibodies for FMDV research. Catalog numbers are clickable links to the product pages.
Protein
| Catalog No. |
Product name |
| VK500132 |
Recombinant FMDV (SAT-1) VP1 Protein, N-His |
| VK164012 |
Recombinant FMDV (SAT-1) VP2 Protein, N-His |
| VK164022 |
Recombinant FMDV (SAT-1) VP3 Protein, N-His |
| VK164032 |
Recombinant FMDV (SAT-1) VP4 Protein, N-His |
| VK164042 |
Recombinant FMDV (SAT-1) Protease 3C, N-His |
| VK500122 |
Recombinant FMDV VP0 Protein, N-His-KSI |
| VK500052 |
Recombinant FMDV Lpro Protein, N-His |
| VK500062 |
Recombinant FMDV VP2 Protein, N-His |
| VK500072 |
Recombinant FMDV VP3 Protein, N-His |
| VK500082 |
Recombinant FMDV VP1 Protein, N-His |
| VK500092 |
Recombinant FMDV Protein 3A, N-His |
| VK500102 |
Recombinant FMDV Protein 2C, N-His |
| VK500112 |
Recombinant FMDV Protease 3C Protein, N-His |
| VK500012 |
Recombinant FMDV Capsid protein VP1 Protein, N-His |
| VK500042 |
Recombinant FMDV VP3 Protein, N-His |
| VK500032 |
Recombinant FMDV VP1 Protein, N-His |
| VK500022 |
Recombinant FMDV VP0 Protein, N-His |
Click to show remaining rows
Antibody
| Catalog No. |
Product name |
| VK500134 |
Anti-FMDV (SAT-1) VP1 Polyclonal Antibody |
| VK164014 |
Anti-FMDV (SAT-1) VP2 Polyclonal Antibody |
| VK164024 |
Anti-FMDV (SAT-1) VP3 Polyclonal Antibody |
| VK164034 |
Anti-FMDV (SAT-1) VP4 Polyclonal Antibody |
| VK164044 |
Anti-FMDV (SAT-1) Protease 3C Polyclonal Antibody |
| VK500124 |
Anti-FMDV VP0 Polyclonal Antibody |
| VK500014 |
Anti-FMDV Capsid protein VP1 Polyclonal Antibody |
| VK500013 |
Anti-FMDV Capsid protein VP1 Antibody (SD6) |
| VK500023 |
Anti-FMDV Capsid protein VP1 Antibody (4C4) |
| VK500033 |
Anti-FMDV Capsid protein VP1 Antibody (4A2) |
| VK500043 |
Anti-FMDV Capsid protein VP1 Antibody (1E12#) |
| VK500053 |
Anti-FMDV Genome polyprotein Antibody (54#) |
| VK500063 |
Anti-FMDV Genome polyprotein Antibody (201#) |
| VK657013 |
Anti-Foot-and-mouth disease virus/FMDV 3ABC/3B Antibody (FM27) |
| VK500073 |
Anti-FMDV Genome polyprotein Antibody (3B3A5) |
| PTX19268 |
Anti-FMDV Capsid protein VP1 Antibody (SD6) |
| PTX19269 |
Anti-FMDV Capsid protein VP1 Antibody (4C4) |
| PTX19270 |
Anti-FMDV Capsid protein VP1 Antibody (4A2) |
| PTX19271 |
Anti-FMDV Capsid protein VP1 Antibody (1E12#) |
| VK500024 |
Anti-FMDV VP0 Polyclonal Antibody |
| VK500034 |
Anti-FMDV VP1 Polyclonal Antibody |
| VK500044 |
Anti-FMDV VP3 Polyclonal Antibody |
| VK500064 |
Anti-FMDV VP2 Polyclonal Antibody |
| VK500074 |
Anti-FMDV VP3 Polyclonal Antibody |
| VK500084 |
Anti-FMDV VP1 Polyclonal Antibody |
Click to show remaining rows
Need AIH research tools?
abinScience offers factory-direct recombinant proteins, antibodies, and ELISA kits for autoimmune hepatitis research, supporting bulk inquiries, custom specifications, and technical consultation.
Email: info@abinscience.com | Tel: +86-027-65523339
Reference:
1. Dong H, Liu P, Bai M, et al. Structural and molecular basis for foot-and-mouth disease virus neutralization by two potent protective antibodies. Protein Cell. 2022;13(6):446-453. doi:10.1007/s13238-021-00828-9
2. Park JY, Lee HM, Kang KJ, et al. Development and immunogenicity of adenoviral Fc-fused FMDV virus-like particle vaccine in swine. Vet Q. 2025;45(1):2564443. doi:10.1080/01652176.2025.2564443
3. Kotecha A, Seago J, Scott K, et al. Structure-based energetics of protein interfaces guides foot-and-mouth disease virus vaccine design. Nat Struct Mol Biol. 2015;22(10):788-794. doi:10.1038/nsmb.3096
4. Porta C, Kotecha A, Burman A, et al. Rational engineering of recombinant picornavirus capsids to produce safe, protective vaccine antigen. PLoS Pathog. 2013;9(3):e1003255. doi:10.1371/journal.ppat.1003255
5. Sultanov AA, Tyulegenov S, Yessembekova GN, et al. The progressive control of foot-and-mouth disease (FMD) in the Republic of Kazakhstan: Successes and challenges. Front Vet Sci. 2023;10:1036121. Published 2023 Apr 17. doi:10.3389/fvets.2023.1036121
6. Li Y, Zeng W, Niu X, et al. Foot-and-mouth disease virus-like particle vaccine incorporating dominant T and B cell epitopes: enhanced immune response in piglets with CD154 molecules. Front Vet Sci. 2025;12:1540102. Published 2025 Feb 19. doi:10.3389/fvets.2025.1540102
7. Gao Y, Sun SQ, Guo HC. Biological function of Foot-and-mouth disease virus non-structural proteins and non-coding elements. Virol J. 2016;13:107. Published 2016 Jun 22. doi:10.1186/s12985-016-0561-z
8. Stenfeldt C, Eschbaumer M, Humphreys J, Medina GN, Arzt J. The pathogenesis of foot-and-mouth disease virus: current understandings and knowledge gaps. Vet Res. 2025;56(1):119. Published 2025 Jun 16. doi:10.1186/s13567-025-01545-5
9. Jiang S, Yang S, Zhang X, et al. Evolutionary and structural insights into VP1 epitopes of representative SAT-type FMDV strains: implications for candidate vaccine selection. Vet Res. 2025;56(1):227. Published 2025 Dec 15. doi:10.1186/s13567-025-01643-4
10. Elrashedy A, Mousa W, Nayel M, et al. Systematic review and meta-analysis of the effectiveness of polypeptide, virus-like particles, and viral vector vaccines for foot-and-mouth disease (2020-2025). Sci Rep. 2025;15(1):39370. Published 2025 Nov 10. doi:10.1038/s41598-025-24078-5
11. Yu SC, Lee IK, Kong HS, et al. Foot-and-Mouth Disease Virus-like Particles Produced in E. coli as Potential Antigens for a Novel Vaccine. Vet Sci. 2025;12(6):539. Published 2025 Jun 2. doi:10.3390/vetsci12060539
12. Reeve R, Borley DW, Maree FF, Upadhyaya S, Lukhwareni A, Esterhuysen JJ, Harvey WT, Blignaut B, Fry EE, Parida S, Paton DJ, Mahapatra M. Tracking the Antigenic Evolution of Foot-and-Mouth Disease Virus. PLoS One. 2016 Jul 22;11(7):e0159360. doi: 10.1371/journal.pone.0159360 . PMID: 27448206; PMCID: PMC4957747.

 中文
中文 English
English 한국어
한국어 日本語
日本語 Español
Español Français
Français Русский
Русский


