Coxiella burnetii Research Advances and Scientific Tools: Insights from Australia's 2025 Q Fever Surge
Release date:
2026-05-26 View count: 3
In April 2026, the Australian Department of Health and Aged Care released a landmark report documenting 912 confirmed Q fever cases in 2025, representing a 52% increase above the 10-year national average.
The causative agent of Q fever is the bacterium Coxiella burnetii. Classified as a CDC Category B priority biothreat agent, C. burnetii possesses extraordinary environmental resilience and efficient aerosol transmission. While acute Q fever can be effectively treated with tetracycline antibiotics, the 1–5% risk of progression to chronic, life-threatening endocarditis remains a critical unmet clinical need.
Beyond these clinical challenges, C. burnetii continues to intrigue basic researchers. Despite the availability of effective drugs for acute infection, many unresolved mysteries remain — including its intracellular parasitism mechanisms, immune evasion strategies, and the molecular basis of chronic infection. Unraveling these puzzles is essential for addressing the lingering threat of this resilient pathogen.
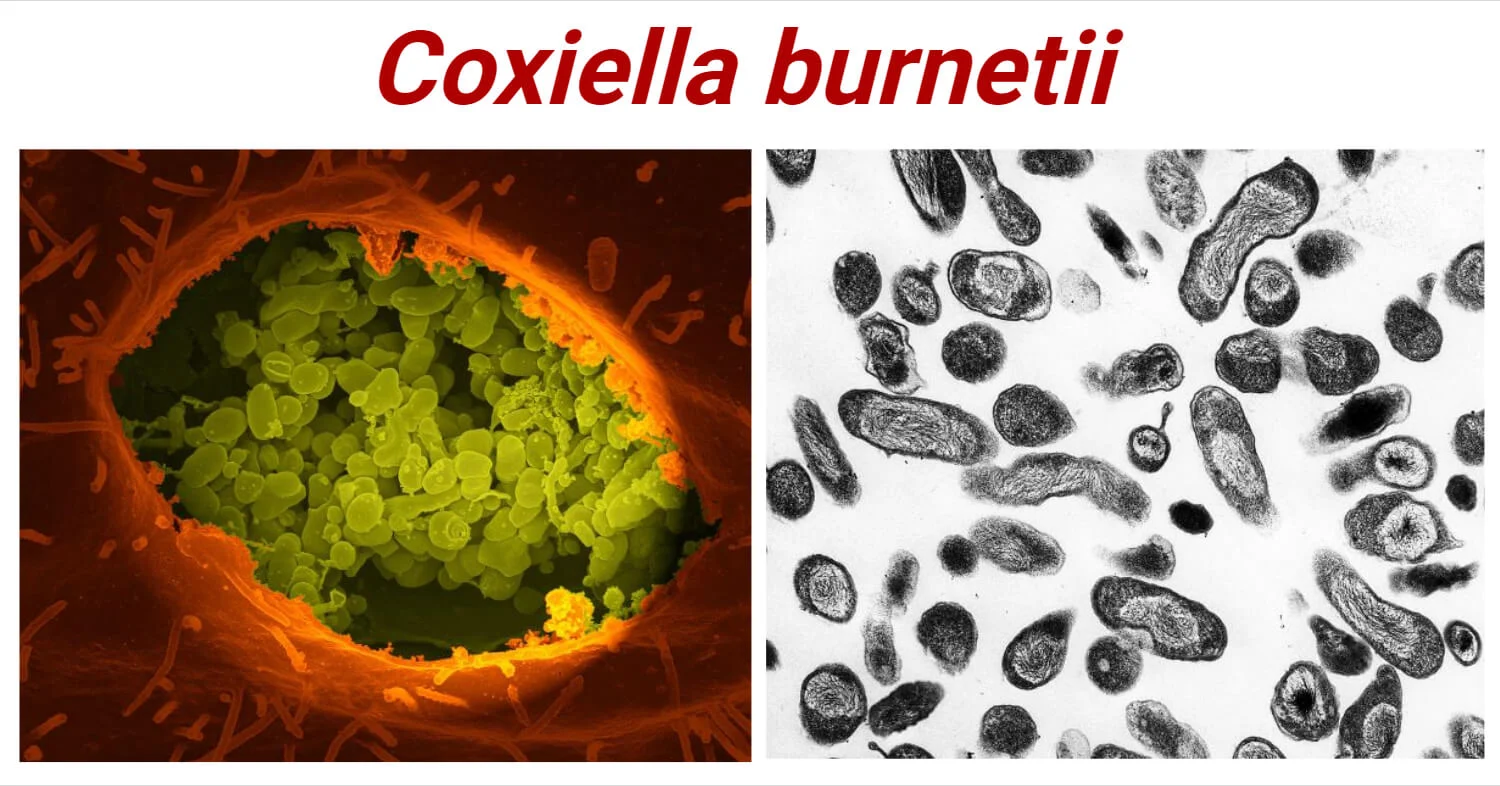
Figure 1: Coxiella burnetii
(Source: CDC Public Health Image Library (PHIL))
1. Biological Characteristics and Taxonomic Status of Coxiella burnetii
1.1 Taxonomic Reclassification
Traditionally, C. burnetii was classified in the order Rickettsiales due to morphological similarities with rickettsiae. However, modern molecular biology studies have shown that its genomic characteristics are more closely related to the class Gammaproteobacteria. It is now formally classified as: Phylum Proteobacteria, Class Gammaproteobacteria, Order Legionellales, Family Coxiellaceae, Genus Coxiella, and is currently the only confirmed species in this genus. This taxonomic adjustment is of great significance for understanding its evolutionary relationships and biological characteristics.
1.2 Unique Biphasic Developmental Cycle
The most prominent biological feature of C. burnetii is its biphasic developmental cycle, in which two morphologically and functionally distinct cell types alternate within host cells:
• Large Cell Variant (LCV): Approximately 0.5–2.0 μm in size, metabolically active and replicative, primarily residing within the parasitophorous vacuoles (PVs) of host cells
• Small Cell Variant (SCV): Approximately 0.2–0.5 μm in size, metabolically dormant with a spore-like structure, exhibiting extreme resistance to desiccation, high temperatures, ultraviolet radiation, and common disinfectants, and can survive in the external environment for months
This biphasic developmental strategy enables C. burnetii to replicate efficiently within host cells while persisting in harsh environments, providing the biological basis for its aerosol transmission.
1.3 Intracellular Parasitism Characteristics
C. burnetii is an obligate intracellular bacterium that primarily infects the mononuclear phagocyte system. Unlike most intracellular bacteria, it not only resists the bactericidal effects of phagolysosomes but also proliferates abundantly in the acidic phagolysosomal environment at pH 4.5–5.0. It remodels the host cell membrane system to form a unique parasitophorous vacuole (PV), which possesses lysosomal characteristics but is hijacked by C. burnetii for its own replication.
2. Pathogenic Mechanisms and Immune Evasion Strategies
2.1 Major Virulence Factors
2.1.1 Type IV Secretion System (T4SS)
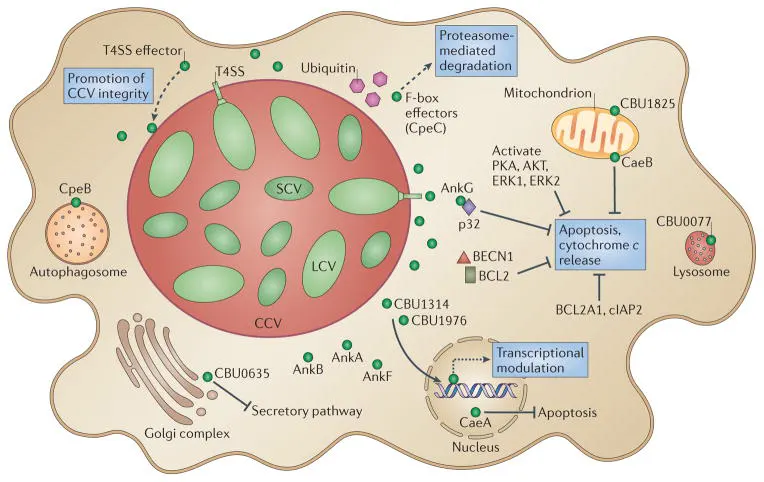
C. burnetii possesses a complete Dot/Icm type IVB secretion system, which is its most critical virulence factor. This system can secrete more than 100 effector proteins into host cells to regulate various physiological processes of the host, including:
• Inhibition of host cell apoptosis
• Promotion of parasitophorous vacuole formation and maturation
• Regulation of host cell autophagy
• Interference with host immune signaling pathways
Figure 2. Roles of type IV secretion system effectors during Coxiella burnetii intracellular infection
(Source: DOI: 10.1038/nrmicro3049)
2.1.2 Lipopolysaccharide (LPS) Phase Variation
C. burnetii exists in two antigenic variants: Phase I and Phase II:
• Phase I: Possesses a complete LPS structure, is highly virulent, and represents wild-type strains isolated from naturally infected animals or patients
• Phase II: Has a truncated LPS structure lacking the O-specific polysaccharide side chain, exhibits attenuated virulence, and is a variant obtained after continuous laboratory passage
This phase variation is of great significance in serological diagnosis: patients with acute Q fever primarily produce antibodies against Phase II antigens, while patients with chronic Q fever are characterized by high titers of Phase I antibodies.
2.1.3 Outer Membrane Protein Family (ompA/ompP1)
• ompA (CBU0307): As the major outer membrane protein, it mediates bacterial adhesion to and invasion of host cells, is a key protein involved in parasitophorous vacuole formation, and exhibits strong immunogenicity, making it an important target for serological diagnosis
• ompP1: An outer membrane porin that regulates material exchange between bacteria and host cells, plays a role in bacterial adaptation to the acidic phagolysosomal environment, and its expression level is positively correlated with bacterial virulence
2.1.4 Heat Shock Protein (groEL)
• Belongs to the HSP60 family, is highly expressed when bacteria respond to intracellular stress environments (such as acidity and oxidative stress), and maintains the correct folding of other virulence proteins
• Possesses cross-immunogenicity, can activate host T-cell immune responses, and is a potential target for chronic Q fever diagnosis and vaccine development
2.1.5 Metabolism-Related Proteins (sucB/icd/gcvT)
• sucB: Participates in the tricarboxylic acid cycle, regulates bacterial energy metabolism, and maintains bacterial survival in nutrient-limited intracellular environments
• icd: Isocitrate dehydrogenase that mediates carbon metabolism and redox balance, affecting the synthesis of bacterial virulence factors
• gcvT: Participates in the glycine cleavage pathway, provides key metabolic intermediates for bacteria, and is an important metabolic node for intracellular replication
2.2 Immune Evasion Mechanisms
C. burnetii has evolved multiple sophisticated immune evasion strategies:
1. Mediates host cell invasion via ompA/ompP1, rapidly enters phagolysosomes, and establishes parasitophorous vacuoles
2. Upregulates groEL expression to resist host stress environments and maintain virulence protein function
3. Regulates the expression of metabolism-related proteins (sucB/icd/gcvT) to adapt to the intracellular nutritional environment
4. Utilizes LPS phase variation to reduce recognition efficiency by the host immune system and achieve persistent infection
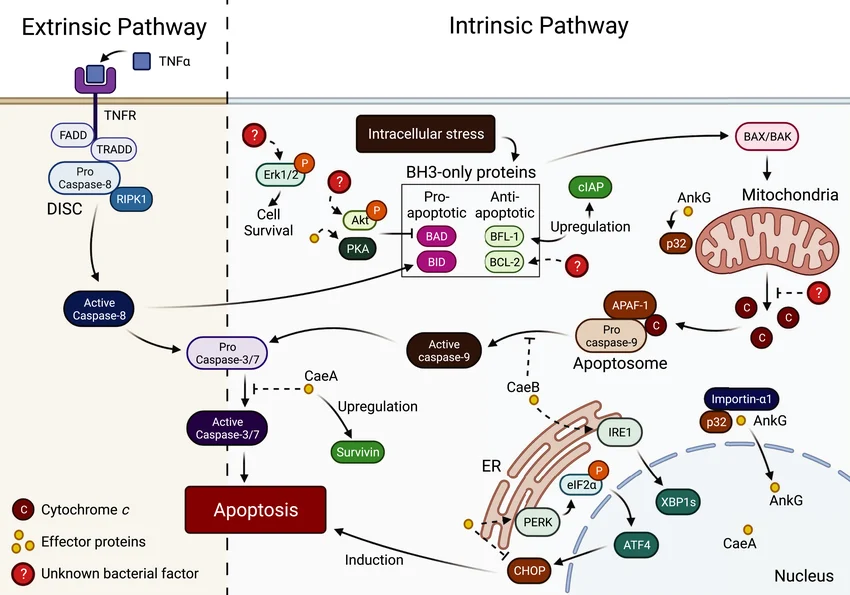
Figure 3. Interactions between Coxiella burnetii and apoptotic pathways
(Source: DOI: 10.1111/mmi.14878)
3. Key Research Hotspots and Challenges
3.1 Research on Chronic Infection Mechanisms
Approximately 1–5% of patients with acute Q fever develop chronic infection, which primarily manifests as endocarditis and vascular infections. The treatment cycle lasts 18–24 months, and the mortality rate can reach 10–25% without timely and standardized treatment. The mechanisms underlying chronic infection are not yet fully understood and may be related to the following factors:
• Impaired host immune function
• Bacterial persistence
• Specific bacterial genotypes
• Host genetic factors
Elucidating the molecular mechanisms of chronic infection is of great significance for developing new therapeutic strategies and vaccines.
3.2 Vaccine Development
Currently, only the Australian-produced Q-Vax® vaccine is commercially available worldwide. This vaccine is prepared from inactivated Phase I C. burnetii and has a protection rate of over 95%. However, its application is limited due to potential severe local and systemic reactions and its unsuitability for previously infected individuals.
Research directions for next-generation vaccines include:
• Subunit vaccines: such as recombinant Com1 protein and HSP proteins
• Live-attenuated vaccines: generated by knocking out virulence genes via genetic engineering
• DNA vaccines: plasmid DNA encoding protective antigens
3.3 Biosafety and Biodefense
Due to its aerosol transmission capability, extremely low infectious dose, and strong environmental resistance, C. burnetii is considered a potential biological warfare agent and bioterrorism agent. Therefore, research on rapid detection technologies, therapeutic drugs, and vaccines for C. burnetii is of strategic importance for national biosafety.
abinScience Products for Coxiella burnetii Research
To meet the research needs on C. burnetii, abinScience offers a comprehensive portfolio of high-quality Coxiella burnetii-related recombinant proteins and antibodies. All products are for research use only.
[1] Osbron, C. A., & Goodman, A. G. (2022). To die or not to die: Programmed cell death responses and their interactions with Coxiella burnetii infection. Molecular microbiology, 117(4), 717–736. https://doi.org/10.1111/mmi.14878
[2] van Schaik, E. J., Chen, C., Mertens, K., Weber, M. M., & Samuel, J. E. (2013). Molecular pathogenesis of the obligate intracellular bacterium Coxiella burnetii. Nature reviews. Microbiology, 11(8), 561–573. https://doi.org/10.1038/nrmicro3049
[3] Beare, P. A., Jeffrey, B. M., Long, C. M., Martens, C. M., & Heinzen, R. A. (2018). Genetic mechanisms of Coxiella burnetii lipopolysaccharide phase variation. PLoS pathogens, 14(3), e1006922. https://doi.org/10.1371/journal.ppat.1006922
[4] Vranakis, I., Mathioudaki, E., Kokkini, S., & Psaroulaki, A. (2019). Com1 as a Promising Protein for the Differential Diagnosis of the Two Forms of Q Fever. Pathogens (Basel, Switzerland), 8(4), 242. https://doi.org/10.3390/pathogens8040242
[5] Wang, T., Wang, C., Li, C., & Song, L. (2023). The intricate dance: host autophagy and Coxiella burnetii infection. Frontiers in microbiology, 14, 1281303. https://doi.org/10.3389/fmicb.2023.1281303
[6] Bauer, B. U., Knittler, M. R., Andrack, J., Berens, C., Campe, A., Christiansen, B., Fasemore, A. M., Fischer, S. F., Ganter, M., Körner, S., Makert, G. R., Matthiesen, S., Mertens-Scholz, K., Rinkel, S., Runge, M., Schulze-Luehrmann, J., Ulbert, S., Winter, F., Frangoulidis, D., & Lührmann, A. (2023). Interdisciplinary studies on Coxiella burnetii: From molecular to cellular, to host, to one health research. International journal of medical microbiology : IJMM, 313(6), 151590. https://doi.org/10.1016/j.ijmm.2023.151590

 中文
中文 English
English 한국어
한국어 日本語
日本語 Español
Español Français
Français Русский
Русский